OUCH! THAT HURTS!
PAIN PERCEPTION IN ANIMALS

There is no getting around it: hunters often inflict pain on their quarry. Ideally a hunt is carried to completion in a manner that causes the least distress to the game, and death comes so rapidly the animal is almost unaware it's happened. But regrettably, not every shot is perfectly placed, not every animal is killed instantaneously, and sometimes we cause more pain than we intend. Anyone who hunts needs a basic understanding of the mechanism of pain perception, not only to avoid inflicting it if at all possible, but to grasp the full importance of ethical hunting technique.
It's sometimes said that, "...animals don't feel pain the way we do." This is wrong. Anyone who's stepped on a dog's foot is quite well aware that higher beings (which certainly includes all mammals and birds, and probably almost all vertebrates) have a similar reaction to such stimuli. If a man drops a rock on his toe we say he "screeches in pain," but in animals, scientists hedge their bets (and avoid the cardinal sin of attributing human emotions to animals) by using the term "exhibits a pain-like response," for the same noise, elicited by the same stimulus. This is quibbling: an animal clearly is capable of feeling "pain" in the same way we do.
So exactly what is he feeling? Pain research and the literature on pain are extensive, but for a variety of reasons, this is still an area of physiology that's not very well understood. Part of the problem is that one can't ethically conduct experiments in serious pain infliction using human subjects; and when animal models are substituted, there's no way to be absolutely certain that the model is 100% accurate. Also, the vast bulk of research is on chronic pain in humans, typically pain associated with diseases like arthritis. This is important but may or may not be the same thing as acute traumatic pain, the kind associated with gunshot wounds. Studies on the latter are fewer, almost always involve animal models, and usually are associated with spinal cord injuries. Nevertheless, some things about acute and chronic pain perception are well understood in general terms, if not in detail.

First, it's important to realize pain, like any other sensation (such as heat, cold, touch, smell, or even color) is "centrally" perceived. That is to say, the actual sensation of pain is not located in the injured area, but in the central nervous system, i.e., the brain and spinal cord. The sensory structures in the periphery are signal transducers and routers; they send information to the brain, and it's there that the stimulus is determined to be "painful" or not. The site of action of anesthetics and chemical pain-killers is usually the central nervous system, not the site where the pain sensation is generated.
Pain perception is the province of innumerable small "naked" nerve fibers, collectively termed "nociceptors" (from the Latin noceo, to hurt or injure). These "free nerve endings" are distributed everywhere throughout an animal's body, usually in close association with blood vessels and certain cell types to form functional "pain units." Nociceptors, like any other sensory fiber, respond to stimuli by sending an electrical signal into the central nervous system (CNS), where its arrival is duly noted and processed by higher-level neurons. In other words, the nociceptor is a sort of switch. ON = pain, OFF = no pain. More accurately, it functions in somewhat the manner of a rheostat, because its output usually varies with the degree of stimulation. As the stimulus increases in intensity the nociceptor fiber fires off its signal more rapidly, sending signals to the CNS at shorter intervals; the brain then processes this information as greater pain.
At the point in the central nervous system where the signal arrives, the nociceptor fiber releases transmitter chemicals whose presence is perceived by a second set of "integrator" neurons. These integrators have options about what to do. They can relay the signal to the coordinating centers in the brain who determine what to do about the incoming sensation of pain, but they also can respond to a high-frequency signal on their own, without reference to Higher Headquarters, at least for a short time. A good deal of the immediate processing of the signals doesn't even reach the brain, actually. Much of it is handled by lower level centers in the spinal cord.
The integrator neurons of the spinal cord are believed in most cases to have an inhibitory effect on the signal. They monitor the rate of transmission of information from the periphery and can unilaterally initiate a first response that temporarily delays the relay of the pain sensation to the brain. The logic behind this sort of lower-level processing is quite simple: pain, especially acute pain, is a signal to the animal that something is wrong and needs to be dealt with now. By routing the signal through the shortest possible pathway and not sending it up to the brain and back down every time, the immediate response is much faster. Naturally the brain has to be "told" sooner or later, but if immediate action is warranted the integration capability of the spinal cord has that "authority."
 Here's an example: if you pat a horse on the rump lightly he doesn't mind. But if you hit a horse's rump with a stone from a slingshot, he'll immediately respond to this sharp stimulus by running away. If you could measure the rate of transmission of the signal in various parts of the nervous system you could easily show that his legs started moving before the brain was actually aware of what had happened. The sensory-motor "loop" of signals that initiates the running response is routed through the spinal cord alone. The signal from the periphery indicating "IMPACT ON RUMP" first gets his muscles moving, and it gets to the command-and-control regions of the cerebral cortex second because it has farther to go and because that sharp impact just might have been something dangerous. Even in the case of a non-threatening "injury" like this, the system responds immediately, because, well...you never know, it might have been a bullet after all. In most cases a wounded animal's first impulse is to run; regardless of the severity of the wound, he is going to get out of the area as fast as possible and look into the details of how badly he's been hurt later. The survival value of this arrangement is obvious. The ones that run away may survive, the ones that don't or can't will almost surely die.
Here's an example: if you pat a horse on the rump lightly he doesn't mind. But if you hit a horse's rump with a stone from a slingshot, he'll immediately respond to this sharp stimulus by running away. If you could measure the rate of transmission of the signal in various parts of the nervous system you could easily show that his legs started moving before the brain was actually aware of what had happened. The sensory-motor "loop" of signals that initiates the running response is routed through the spinal cord alone. The signal from the periphery indicating "IMPACT ON RUMP" first gets his muscles moving, and it gets to the command-and-control regions of the cerebral cortex second because it has farther to go and because that sharp impact just might have been something dangerous. Even in the case of a non-threatening "injury" like this, the system responds immediately, because, well...you never know, it might have been a bullet after all. In most cases a wounded animal's first impulse is to run; regardless of the severity of the wound, he is going to get out of the area as fast as possible and look into the details of how badly he's been hurt later. The survival value of this arrangement is obvious. The ones that run away may survive, the ones that don't or can't will almost surely die.
But even when a traumatic injury is very bad, there is considerable evidence that pain isn't felt immediately, certainly in most cases not nearly as severely as it may be felt later. As one review article phrases it:
"Events occurring in the periphery and in the [spinal cord] can cause a dissociation of pain perception from the presence or degree of actual tissue injury." (Pain: Neuroanatomy, Chemical Mediators, and Clinical Implications, May 2000 issue of AACN Clinical Issues, p 168.)
It's the inhibitory effect of the central nervous system integrator neurons that's primarily responsible for this delayed perception of pain (not, as is commonly assumed, the adrenalin released in the "fight or flight" response). CNS activity is usually very rapid in this respect: again, the survival value of a temporary and nearly instantaneous shutdown of pain perception is clear.
Another well known response to trauma is the release of materials that have many of the same chemical actions as opiate used clinically as pain killers. These are released from special neurons of the CNS, triggered by the incoming signal relayed to them by integrator neurons. These "endogenous opioids" can completely block the central perception of pain for quite some time: certainly for seconds, usually for several minutes or even longer. The integrator neurons of the spinal cord tell other neurons in the pons, the midbrain, and the medulla what's happened. These neurons send fibers back to the spinal cord, where they secrete their opioids. The opioids modulate the activity of the integrator neurons to further inhibit nociceptor input and extend the immediate pain-deadening response for some additional time.
The phenomenon is well known in humans. It's quite usual for people who have suffered severe traumatic injuries in accidents to report they "felt no pain" initially, often not until the next day, if even then. While endogenous opioid action isn't as fast as the immediate suppression mechanism described above, it's a very valuable and important effect. The nearly instantaneous neuronal suppression of pain perception gives the animal a chance to start running; while he's doing that, production of endogenous opioids is ramped up, which confers the ability to hang on until he reaches a safe place. Delayed perception of pain by production of endogenous opioids amounts to a form of "self anesthesia." This mechanism explains why animals with very serious trauma are often capable of going long distances, seemingly unaffected by their wounds. They are affected, but for some considerable period of time, they're producing internal pain-killers that mitigate the pain of the injury, allowing them to temporarily ignore everything except the need to escape.
Interestingly, the opioid-secreting regions of the brain have a very long evolutionary history, going back to the earliest origins of vertebrates; the reactions they control are typically automatic functions vital to individual survival. "Automating" a delayed response to painful trauma is certainly evolutionarily favorable. As with any other metabolic reaction, there's a lot of variation among individuals, but over eons of time it can be expected that Mother Nature will select for survival those individuals who are capable of rapid, high-level production of opioids and hence a better chance of escaping predators.
There's still another mechanism, at the level of the nociceptor neurons themselves, that can confer pain resistance not just in the short term, but permanently. Almost everyone knows someone who has "a high pain tolerance," an individual whose ability to tolerate discomfort is such that he willingly undergoes dental procedures or even minor surgery without anesthetics. Such people actually may refuse anesthesia when being treated for relatively serious cuts or burns, saying "It doesn't hurt that much." While this may be dismissed by a campmate or worried spouse as mere bravado and machismo, there is sound reason from recent research to believe that a high pain threshold is an inherited trait.
The membranes of nociceptor neurons in certain individuals may have fewer than the normal number of receptor sites for pain sensation. Fewer receptor molecules mean those nociceptors require a much higher level of stimulus to fire off the initial signal that tells the CNS "I'm hurting!" A group of neuroscientists in California, Maryland, and
Now, this is a very interesting finding. Demonstrating the existence of at least one receptor protein dedicated to receiving specific types of stimuli implies very strongly that similar receptor molecules exist for all classes of potentially painful sensations. If nociceptor neurons are in fact stimulus-specific, a genetic absence of the ability to make one type of nociceptor (or modification of its response properties) implies that an otherwise "painful" stimulus may simply not exist for that individual. This model easily accounts for the variations among individuals of a species in pain tolerance. Any animal whose production of receptor proteins for a given stimulus is low level will be "pain tolerant" for that (and perhaps other) stimuli.
 The stoicism of animals—and of humans, too—who've been injured is the stuff of legend. Many cultures have rites for entry into adulthood that involve tolerating pain. Most hunters have encountered game animals that have survived despite sustaining serious wounds. The ability to cope with significant injuries without obvious discomfort is something almost any dog owner is familiar with. (I once had a dog who had a very large tumor removed from his back; he was half-flayed in the surgery but never showed the least sign of pain upon recovering from anesthesia. I'd have died from shock had someone done that to me, I can tell you.)
The stoicism of animals—and of humans, too—who've been injured is the stuff of legend. Many cultures have rites for entry into adulthood that involve tolerating pain. Most hunters have encountered game animals that have survived despite sustaining serious wounds. The ability to cope with significant injuries without obvious discomfort is something almost any dog owner is familiar with. (I once had a dog who had a very large tumor removed from his back; he was half-flayed in the surgery but never showed the least sign of pain upon recovering from anesthesia. I'd have died from shock had someone done that to me, I can tell you.)
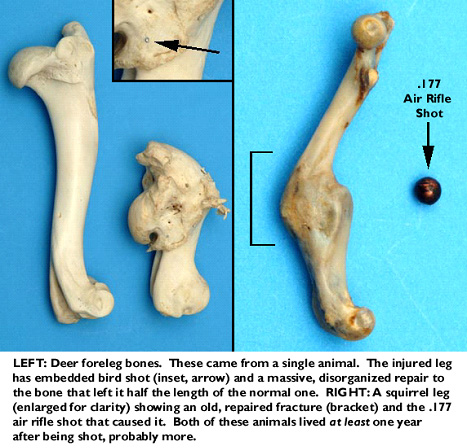
I have included two such examples: one is the shattered humerus (upper foreleg bone) of a white-tailed deer. This deer was shot by some imbecile with a load of small birdshot at close range, in an act of wanton sadism. This deer healed (though with loss of function) and lived for at least a year afterwards, probably longer: the bone has been extensively remodeled and repaired and it's clearly that of a mature animal. The second example is the femur of a tree squirrel. Again, this leg bone has a very old and well-repaired fracture, caused by an air rifle bullet, which I found embedded in the leg muscle close to the injury it caused. Nor are such phenomena confined to limbs: I have handled a hippo skull that had not one, but three badly corroded .303 military bullets embedded in it. The hippo had been killed by a hunter who took it with a single heart-lung shot, and who found the bullets when the skull was cleaned for display. While there's no doubt that the hippo had a phenomenal headache for a while, he lived. These animals, all victims of callousness, inadequately-powered weapons, or bad marksmanship, certainly felt pain; but the fact that they were equally clearly able to make good their escape and hole up somewhere until the injury was dealt with indicates that their tolerance for it must have been very high indeed. Probably that pain tolerance was passed along to their offspring, too.
So, then, what does all this mean to the hunter? How does understanding the mechanisms of pain affect the decisions that must be made to shoot or not, and the obligation to kill quickly and cleanly? To me, the situation is quite clear.
First, there is no question at all that animals can feel pain just as we do: hence to my mind taking a shot when you seriously doubt it will be immediately effective in putting the quarry down for keeps should be avoided. To do otherwise is cruel because it greatly increases the odds that the animal will suffer needlessly. The fact that wild animals are more tolerant of pain that the somewhat soft humans who hunt them is irrelevant to the issue: if an animal is allowed to escape with a wound, fairly soon thereafter it will be hurting very badly, as badly as any human would be with the same injury. While nothing is an absolute certainty in the field, no hunter can absolve himself of the responsibility to do all he can to make every shot effective, and to effect a kill instantly if possible. A clean kill or a clean miss is the only ethical approach.

That said, I think it may be of some importance to hunters to know that for some discrete but unpredictable period of time after being shot the animal is not in great pain, though he may eventually be. The typical situation of an animal downed but not immediately killed is one in which the various mechanisms of pain suppression discussed here are operative. While there is no way to tell exactly how long the numbing effect of inhibitory neurons and endogenous opioids lasts in any individual case, for a brief while the animal may be frightened and disoriented, but he's not "in pain" in the classic sense. Those invaluable seconds-to-minutes are the time for a quick follow-up shot to see to it that he doesn't make the transition from bewilderment to agony.
It's the ones that get up and run a long way, and worse, the ones whose spoor you lose, that eventually do make that transition. If the injury is severe enough they'll likely die, and it will not be a pleasant death. Anyone who takes to the field with gun in hand has to understand this and accept the reality of what he may do if he's careless, under-gunned, out of practice, or badly trained. Quite aside from the foolishness of spending a great deal of money on a hunt that turns out badly for these reasons, it's ethically unacceptable. The hunter has an obligation to the hunted to do his best to see to it that the latter's mechanism of pain perception is never used.
| HUNTING | GUNS | DOGS |
| FISHING & BOATING | TRIP REPORTS | MISCELLANEOUS ESSAYS |
| CONTRIBUTIONS FROM OTHER WRITERS|
| RECIPES |POLITICS |